Молекулярная биология. В чем заключается теория РНК-мира? Первичный мир рнк
© А.С. Спирин
БИОСИНТЕЗ БЕЛКОВ, МИР РНКИ ПРОИСХОЖДЕНИЕ ЖИЗНИ
А.С. Спирин
Спирин Александр Сергеевич - академик, директор Института белка РАН, член Президиума РАН.
Почти полвека тому назад, в 1953 г., Д. Уотсон и Ф. Крик открыли принцип структурной (молекулярной) организации генного вещества - дезоксирибонуклеиновой кислоты (ДНК) . Структура ДНК дала ключ к механизму точного воспроизведения - редупликации - генного вещества . Так возникла новая наука - молекулярная биология. Была сформулирована так называемая центральная догма молекулярной биологии: ДНК Ю РНК Ю белок. Смысл ее состоит в том, что генетическая информация, записанная в ДНК, реализуется в виде белков, но не непосредственно, а через посредство родственного полимера - рибонуклеиновую кислоту (РНК), и этот путь от нуклеиновых кислот к белкам необратим. Таким образом, ДНК синтезируется на ДНК, обеспечивая собственную редупликацию, то есть воспроизведение исходного генетического материала в поколениях; РНК синтезируется на ДНК, в результате чего происходит переписывание, или транскрипция, генетической информации в форму многочисленных копий РНК; молекулы РНК служат матрицами для синтеза белков - генетическая информация транслируется в форму полипептидных цепей. В специальных случаях РНК может переписываться в форму ДНК ("обратная транскрипция"), а также копироваться в виде РНК (репликация), но белок никогда не может быть матрицей для нуклеиновых кислот (подробнее см. ).
Итак, именно ДНК определяет наследственность организмов, то есть воспроизводящийся в поколениях набор белков и связанных с ними признаков. Биосинтез белка является центральным процессом живой материи, а нуклеиновые кислоты обеспечивают его, с одной стороны, программой, определяющей весь набор и специфику синтезируемых белков, а с другой - механизмом точного воспроизведения этой программы в поколениях. Следовательно, происхождение жизни в ее современной клеточной форме сводится к возникновению механизма наследуемого биосинтеза белков.
БИОСИНТЕЗ БЕЛКОВ
Центральная догма молекулярной биологии постулирует лишь путь передачи генетической информации от нуклеиновых кислот к белкам и, следовательно, к свойствам и признакам живого организма. Изучение механизмов реализации этого пути на протяжении десятилетий, последовавших за формулировкой центральной догмы, вскрыло гораздо более разнообразные функции РНК, чем быть только переносчиком информации от генов (ДНК) к белкам и служить матрицей для синтеза белков.
На рис. 1 представлена общая схема биосинтеза белка в клетке.
РНК-посредник
(messenger RNA, матричная РНК, мРНК), кодирующая белки, о
которой и шла речь выше, - это лишь один из трех главных классов клеточных
РНК. Основную их массу (около 80%) составляет другой класс РНК - рибосомные
РНК,
которые образуют структурный каркас и функциональные центры
универсальных белок-синтезирующих частиц - рибосом. Именно рибосомные РНК
ответственны - как в структурном, так и в функциональном отношении - за
формирование ультрамикроскопических молекулярных машин, называемых рибосомами.
Рибосомы воспринимают генетическую информацию в виде молекул мРНК и, будучи
запрограммированы последними, делают белки в точном соответствии с данной
программой.
Однако, чтобы синтезировать белки, одной только информации или программы недостаточно - нужен еще и материал, из которого их можно делать. Поток материала для синтеза белков идет в рибосомы через посредство третьего класса клеточных РНК - РНК-переносчиков (transfer RNA, транспортные РНК, тРНК). Они ковалентно связывают - акцептируют - аминокислоты, которые служат строительным материалом для беЛков, и в виде аминоацил-тРНК поступают в рибосомы. В рибосомах аминоацил-тРНК взаимодействуют с кодонами - трехнуклеотидными комбинациями - мРНК, в результате чего и происходит декодирование кодонов в процессе трансляции.
РИБОНУКЛЕИНОВЫЕ КИСЛОТЫ
Итак, перед нами набор главных клеточных РНК, определяющих основной процесс современной живой материи - биосинтез белка. Это мРНК, рибосомные РНК и тРНК. РНК синтезируются на ДНК с помощью ферментов - РНК-полимераз, осуществляющих транскрипцию - переписывание определенных участков (линейных отрезков) двутяжевой ДНК в форму однотяжевой РНК. Участки ДНК, кодирующие клеточные белки, переписываются в виде мРНК, тогда как для синтеза многочисленных копий рибосомной РНК и тРНК имеются специальные участки клеточного генома, с которых идет интенсивное переписывание без последующей трансляции в белки.
Химическая структура РНК.
Химически РНК очень похожа на ДНК. Оба вещества - это линейные полимеры
нуклеотидов. Каждый мономер - нуклеотид - представляет собой фосфорилированный
N-гликозид, построенный из остатка пятиуглеродного сахара - пентозы, несущего
фосфатную группу на гидроксильной группе пятого углеродного атома
(сложноэфирная связь) и азотистое основание при первом углеродном атоме
(N-гликозидная связь). Главное химическое различие между ДНК и РНК состоит в
том, что сахарный остаток мономера РНК - это рибоза, а мономера ДНК -
дезоксирибоза, являющаяся производным рибозы, в котором отсутствует
гидроксильная группа при втором углеродном атоме (рис. 2).
 Рис. 2. Химические формулы остатков одного из рибонуклеотидов - уридиловой кислоты (U) и гомологичного ему дезоксирибонуклеотида - тимидиловой кислоты (dT) |
Азотистых оснований и в ДНК, и в РНК четыре вида: два пуриновых - аденин (А) и гуанин (G) -и два пиримидиновых - цитозин (С) и урацил (U) или его метилированное производное тимин (Т). Урацил характерен для мономеров РНК, а тимин - для мономеров ДНК, и это второе различие РНК и ДНК. Мономеры - рибонуклеотиды РНК или дезоксирибонуклеотиды ДНК - образуют полимерную цепь посредством формирования фосфодиэфирных мостиков между сахарными остатками (между пятым и третьим атомами углерода пентозы). Таким образом, полимерная цепь нуклеиновой кислоты - ДНК или РНК - может быть представлена как линейный сахаро-фосфатный остов с азотистыми основаниями в качестве боковых групп. Макромолекулярная структура РНК. Принципиальное макроструктурное различие двух типов нуклеиновых кислот состоит в том, что ДНК - это единая двойная спираль, то есть макромолекула из двух комплементарно связанных полимерных тяжей, спирально закрученных вокруг общей оси (см. [ , ]), а РНК - однотяжевой полимер. В то же время взаимодействия боковых групп - азотистых оснований - друг с другом, а также с фосфатами и гидроксилами сахаро-фосфатного остова приводят к тому, что однотяжевой полимер РНК сворачивается на себя и скручивается в компактную структуру , подобно сворачиванию полипептидной цепи белка в компактную глобулу. Таким способом уникальные нуклеотидные последовательности РНК могут формировать уникальные пространственные структуры. |
Впервые специфическая пространственная структура РНК была продемонстрирована при расшифровке атомной структуры одной из тРНК в 1974 г. [ , ] (рис. 3). Сворачивание полимерной цепи тРНК, состоящей из 76 нуклеотидных мономеров, приводит к формированию очень компактного глобулярного ядра, из которого под прямым углом торчат два выступа. Они представляют собой короткие двойные спирали по типу ДНК, но организованные за счет взаимодействия участков одной и той же цепи РНК. Один из выступов является акцептором аминокислоты и участвует в синтезе полипептидной цепи белка на рибосоме, а другой предназначен для комплементарного взаимодействия с кодирующим триплетом (кодоном) мРНК в той же рибосоме. Только такая структура способна специфически взаимодействовать с белком-ферментом, навешивающим аминокислоту на тРНК, и с рибосомой в процессе трансляции, то есть специфически "узнаваться" ими.

Рис. 3. Атомная (слева) и скелетная (справа) модели фенилаланиновой тРНК дрожжей
Изучение изолированных рибосомных РНК дало следующий разительный пример формирования компактных специфических структур из еще более длинных линейных полимеров этого типа. Рибосома состоит из двух неравных частей - большой и малой рибосомных субчастиц (субъединиц). Каждая субчастица построена из одной высокополимерной РНК и целого ряда разнообразных рибосомных белков. Длина цепей рибосомных РНК весьма значительна: так, РНК малой субчастицы бактериальной рибосомы содержит более 1500 нуклеотидов, а РНК большой субчастицы - около 3000 нуклеотидов. У млекопитающих, включая человека, эти РНК еще больше - около 1900 нуклеотидов и более 5000 нуклеотидов в малой и большой субчастицах соответственно.
Было показано, что изолированные рибосомные РНК, отделенные от их белковых
партнеров и полученные в чистом виде, сами способны спонтанно сворачиваться в
компактные структуры, по своим размерам и форме похожие на рибосомные
субчастицы ].
Форма большой и малой субчастиц разная, и соответственно различается форма
большой и малой рибосомных РНК (рис. 4). Таким образом, линейные цепи
рибосомной РНК самоорганизуются в специфические пространственные структуры,
определяющие размеры, форму и, по-видимому, внутреннее устройство рибосомных
субчастиц, а следовательно, и всей рибосомы.
Минорные РНК. По мере изучения компонентов живой клетки и отдельных фракций тотальной клеточной РНК выяснялось, что тремя главными видами РНК дело не ограничивается. Оказалось, что в природе существует множество других видов РНК. Это, в первую очередь, так называемые "малые РНК", которые содержат до 300 нуклеотидов, часто с неизвестными функциями. Как правило, они ассоциированы с одним или несколькими белками и представлены в клетке в виде рибонуклеопротеидов - "малых РНП" .
Малые РНК присутствуют во всех отделах клетки, включая цитоплазму, ядро, ядрышко, ми-тохондрии. Большая часть тех малых РНП, функции которых известны, участвует в механизмах посттранскрипционной обработки главных видов РНК (RNA processing) - превращении предшественников мРНК в зрелые мРНК (сплайсинг), редактировании мРНК, биогенезе тРНК, созревании рибосомных РНК. Один из наиболее богато представленных в клетках видов малых РНП (SRP) играет ключевую роль в транспорте синтезируемых белков через клеточную мембрану. Известны виды малых РНК, выполняющих регуляторные функции в трансляции. Специальная малая РНК входит в состав важнейшего фермента, ответственного за поддержание редупликации ДНК в поколениях клеток - теломеразы. Следует сказать, что их молекулярные размеры сопоставимы с размерами клеточных глобулярных белков. Таким образом, постепенно становится ясно, что функционирование живой клетки определяется не только многообразием синтезируемых в ней белков, но и присутствием богатого набора разнообразных РНК, из которых малые РНК в значительной мере имитируют компактность и размеры белков.
Рибозимы. Вся активная жизнь построена на обмене веществ - метаболизме, и все биохимические реакции метаболизма происходят с надлежащими для обеспечения жизни скоростями только благодаря высокоэффективным специфическим катализаторам, созданным эволюцией. На протяжении многих десятилетий биохимики были уверены, что биологический катализ всегда и всюду осуществляется белками, называемыми ферментами , или энзимами. И вот в 1982-1983 гг. было показано, что в природе имеются виды РНК, которые, подобно белкам, обладают высокоспецифической каталитической активностью [ , ]. Такие РНК-катализаторы были названы рибозимами. Представлению об исключительности белков в катализе биохимических реакций пришел конец.
В настоящее время рибосому тоже принято рассматривать как рибозим. Действительно, все имеющиеся экспериментальные данные свидетельствуют о том, что синтез полипептидной цепи белка в рибосоме катализируется рибосомной РНК, а не рибосомными белками. Идентифицирован каталитический участок большой рибосомной РНК, ответственный за катализ реакции транспептидации, посредством которой осуществляется наращивание полипептидной цепи белка в процессе трансляции.
Что касается репликации вирусных ДНК, то ее механизм мало чем отличается от редупликации генетического материала - ДНК - самой клетки. В случае же вирусных РНК реализуются процессы, которые подавлены или вовсе отсутствуют в нормальных клетках, где вся РНК синтезируется только на ДНК как на матрице. При инфекции РНК-содержащими вирусами ситуация может быть двоякой. В одних случаях на вирусной РНК как на матрице синтезируется ДНК ("обратная транскрипция"), а уж на этой ДНК транскрибируются многочисленные копии вирусной РНК. В других, наиболее интересных для нас случаях на вирусной РНК синтезируется комплементарная цепь РНК, которая и служит матрицей для синтеза - репликации - новых копий вирусной РНК. Таким образом при инфекции РНК-содержащими вирусами реализуется принципиальная способность РНК детерминировать воспроизведение своей собственной структуры, как это имеет место у ДНК.
Мультифункциональность РНК. Суммирование и обзор знаний о функциях РНК позволяют говорить о необыкновенной многофункциональности этого полимера в живой природе. Можно дать следующий список основных известных функций РНК.
Генетическая репликативная функция: структурная возможность копирования (репликации) линейных последовательностей нуклеотидов через комплементарные последовательности. Функция реализуется при вирусных инфекциях и аналогична главной функции ДНК в жизнедеятельности клеточных организмов - редупликации генетического материала.В целом РНК предстает перед нами столь удивительным полимером, что, казалось бы, ни времени эволюции Вселенной, ни интеллекта Творца не должно было бы хватить на ее изобретение. Как можно было видеть, РНК способна выполнять функции обоих принципиально важных для жизни полимеров - ДНК и белков. Неудивительно, что перед наукой и встал вопрос: а не могло ли возникновение и самодостаточное существование мира РНК предшествовать появлению жизни в ее современной ДНК-белковой форме?Кодирующая функция: программирование белкового синтеза линейными последовательностями нуклеотидов. Это та же функция, что и у ДНК. И в ДНК, и в РНК одни и те же триплеты нуклеотидов кодируют 20 аминокислот белков, и последовательность триплетов в цепи нуклеиновой кислоты есть программа для последовательной расстановки 20 видов аминокислот в полипептидной цепи белка.
Структурообразующая функция: формирование уникальных трехмерных структур. Компактно свернутые молекулы малых РНК принципиально подобны трехмерным структурам глобулярных белков, а более длинные молекулы РНК могут образовывать и более крупные биологические частицы или их ядра.
Функция узнавания: высокоспецифические пространственные взаимодействия с другими макромолекулами (в том числе белками и другими РНК) и с малыми лигандами. Эта функция, пожалуй, главная у белков. Она основана на способности полимера сворачиваться уникальным образом и формировать специфические трехмерные структуры. Функция узнавания является базой специфического катализа.
Каталитическая функция: специфический катализ химических реакций рибозимами. Данная функция аналогична энзиматической функции белков-ферментов.
ПРОИСХОЖДЕНИЕ ЖИЗНИ
Белково-коацерватная теория Опарина. Пожалуй, первая научная, хорошо продуманная теория происхождения жизни абиогенным путем была предложена биохимиком А.И. Опариным еще в 20-х годах прошлого века [ , ]. Теория базировалась на представлении, что все начиналось с белков, и на возможности в определенных условиях спонтанного химического синтеза мономеров белков - аминокислот - и белковоподобных полимеров (полипептидов) абиогенным путем. Публикация теории стимулировала многочисленные эксперименты в ряде лабораторий мира, показавшие реальность такого синтеза в искусственных условиях. Теория быстро стала общепринятой и необыкновенно популярной.
Основным ее постулатом было то, что спонтанно возникавшие в первичном "бульоне" белковоподобные соединения объединялись" в коацерватные капли - обособленные коллоидные системы (золи), плавающие в более разбавленном водном растворе. Это давало главную предпосылку возникновения организмов - обособление некой биохимической системы от окружающей среды, ее компартментализацию. Так как некоторые белковоподобные соединения коацерватных капель могли обладать каталитической активностью, то появлялась возможность прохождения биохимических реакций синтеза внутри капель - возникало подобие ассимиляции, а значит, роста коацервата с последующим его распадом на части - размножением. Ассимилирующий, растущий и размножающийся делением коацерват рассматривался как прообраз живой клетки (рис. 5).
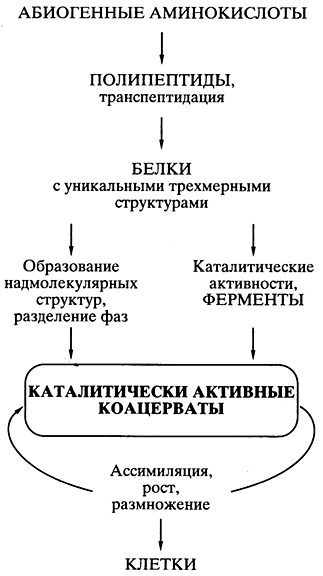
Рис. 5.
Схематическое представление пути
происхождения жизни
согласно белково-коацерватной теории А.И.
Опарина
Все было хорошо продумано и научно обосновано в теории, кроме одной проблемы, на которую долго закрывали глаза почти все специалисты в области происхождения жизни. Если спонтанно, путем случайных безматричных синтезов в коацервате возникали единичные удачные конструкции белковых молекул (например, эффективные катализаторы, обеспечивающие преимущество данному коацервату в росте и размножении), то как они могли копироваться для распространения внутри коацервата, а тем более для передачи коацерватам-потомкам? Теория оказалась неспособной предложить решение проблемы точного воспроизведения - внутри коацервата и в поколениях - единичных, случайно появившихся эффективных белковых структур.
Мир РНК как предшественник современной жизни. Накопление знаний о генетическом коде, нуклеиновых кислотах и биосинтезе белков привело к утверждению принципиально новой идеи о ТОМ, что все начиналось вовсе не с белков, а с РНК [ - ]. Нуклеиновые кислоты являются единственным типом биологических полимеров, макромолекулярная структура которых, благодаря принципу комплементарности при синтезе новых цепей (подробнее см. ), обеспечивает возможность копирования собственной линейной последовательности мономерных звеньев, другими словами, возможность воспроизведения (репликации) полимера, его микроструктуры. Поэтому только нуклеиновые кислоты, но не белки, могут быть генетическим материалом, то есть воспроизводимыми молекулами, повторяющими свою специфическую микроструктуру в поколениях.
По ряду соображений именно РНК, а не ДНК, могла представлять собой первичный генетический материал.
Во-первых, и в химическом синтезе, и в биохимических реакциях рибонуклеотиды предшествуют дезоксирибонуклеотидам; дезоксирибонуклеотиды - продукты модификации рибонуклеотидов (см. рис. 2).
Во-вторых, в самых древних, универсальных процессах жизненного метаболизма широко представлены именно рибонуклеотиды, а не дезоксирибонуклеотиды, включая основные энергетические носители типа рибонуклеозид-полифосфатов (АТФ и т.п.).
В-третьих, репликация РНК может происходить без какого бы то ни было участия ДНК, а механизм редупликации ДНК даже в современном живом мире требует обязательного участия РНК-затравки в инициации синтеза цепи ДНК.
В-четвертых, обладая всеми теми же матричными и генетическими функциями, что и ДНК, РНК способна также к выполнению ряда функций, присущих белкам, включая катализ химических реакций. Таким образом, имеются все основания рассматривать ДНК как более позднее эволюционное приобретение - как модификацию РНК, специализированную для выполнения функции воспроизведения и хранения уникальных копий генов в составе клеточного генома без непосредственного участия в биосинтезе белков.
После того как были открыты каталитически активные РНК, идея первичности
РНК в происхождении жизни получила сильнейший толчок к развитию, и была
сформулирована концепция самодостаточного мира РНК,
предшествовавшего
современной жизни [ ,
].
Возможная схема возникновения мира РНК представлена на рис. 6.
Абиогенный синтез рибонуклеотидов и их ковалентное объединение в олигомеры и полимеры типа РНК могли происходить приблизительно в тех же условиях и в той же химической обстановке, что постулировались для образования аминокислот и полипептидов. Недавно А.Б. Четверин с сотрудниками (Институт белка РАН) экспериментально показали, что по крайней мере некоторые полирибонуклеотиды (РНК) в обычной водной среде способны к спонтанной рекомбинации, то есть обмену отрезками цепи, путем транс-эстерификации . Обмен коротких отрезков цепи на длинные, должен приводить к удлинению полирибонуклеотидов (РНК), а сама подобная рекомбинация способствовать структурному многообразию этих молекул. Среди них могли возникать и каталитически активные молекулы РНК.
Даже крайне редкое появление единичных молекул РНК, которые были способны катализировать полимеризацию рибонуклеотидов или соединение (сплайсинг) олигонуклеотидов на комплементарной цепи как на матрице [ , ], означало становление механизма репликации РНК. Репликация самих РНК-катализаторов (рибозимов) должна была повлечь за собой возникновение самореплицирующихся популяций РНК. Продуцируя свои копии, РНК размножались. Неизбежные ошибки в копировании (мутации) и рекомбинации в самореплицирующихся популяциях РНК создавали все большее разнообразие этого мира. Таким образом, предполагаемый древний мир РНК - это "самодостаточный биологический мир, в котором молекулы РНК функционировали и как генетический материал, и как энзимоподобные катализаторы" .
Возникновение биосинтеза белка. Далее на основе мира РНК должно было происходить становление механизмов биосинтеза белка, появление разнообразных белков с наследуемой структурой и свойствами, компартментализация систем биосинтеза белка и белковых наборов, возможно, в форме коацерватов и эволюция последних в клеточные структуры - живые клетки (см. рис. 6).
Проблема перехода от древнего мира РНК к современному белок-синтезирующему миру - наиболее трудная даже для чисто теоретического решения. Возможность абиогенного синтеза по-липептидов и белковоподобных веществ не помогает в решении проблемы, так как не просматривается никакого конкретного пути, как этот синтез мог бы быть сопряжен с РНК и подпасть под генетический контроль. Генетически контролируемый синтез полипептидов и белков должен был развиваться независимо от первичного абиогенного синтеза, своим путем, на базе уже существовавшего мира РНК. В литературе предложено несколько гипотез происхождения современного механизма биосинтеза белка в мире РНК, но, пожалуй, ни одна из них не может рассматриваться как детально продуманная и безупречная с точки зрения физико-химических возможностей. Представлю свою версию процесса эволюции и специализации РНК, ведущего к возникновению аппарата биосинтеза белка (рис. 7), но и она не претендует на законченность.
Предлагаемая гипотетическая схема содержит два существенных момента, кажущихся принципиальными.
Во-первых, постулируется, что абиогенно синтезируемые олигорибонуклеотиды активно рекомбинировали посредством механизма спонтанной неэнзиматической трансэстерификации , приводя к образованию удлиненных цепей РНК и давая начало их многообразию. Именно этим путем в популяции олигонуклеотидов и полинуклеотидов и могли появиться как каталитически активные виды РНК (рибозимы), так и другие виды РНК со специализированными функциями (см. рис. 7). Более того, неэнзиматическая рекомбинация олигонуклеотидов, комплементарно связывающихся с полинуклеотидной матрицей, могла обеспечить сшивание (сплайсинг) фрагментов, комплементарных этой матрице, в единую цепь. Именно таким способом, а не катализируемой полимеризацией мононуклеотидов, могло осуществляться первичные копирование (размножение) РНК. Разумеется, если появлялись рибозимы, обладавшие полимеразной активностью , то эффективность (точность, скорость и продуктивность) копирования на комплементарной. матрице должна была значительно возрастать.

Рис. 7.
Схема эволюции и специализации молекул РНК
в процессе перехода от древнего мира РНК к современному миру
генетически детерминированного биосинтеза белков
Второй принципиальный момент в моей версии состоит в том, что первичный аппарат биосинтеза белка возник на базе нескольких видов специализированных РНК до появления аппарата энзиматической (полимеразной) репликации генетического материала - РНК и ДНК. Этот первичный аппарат включал каталитически активную прорибосомную РНК, обладавшую пептидил-трансферазной активностью; набор про-тРНК, специфически связывающих аминокислоты или короткие пептиды; другую прорибосомную РНК, способную взаимодействовать одновременно с каталитической прорибосомной РНК, про-мРНК и про-тРНК (см. рис. 7). Такая система уже могла синтезировать полипептидные цепи за счет катализируемой ею реакции транспептидации. Среди прочих каталитически активных белков - первичных ферментов (энзимов) - появились и белки, катализирующие полимеризацию нуклеотидов - репликазы, или НК-полимеразы.
Впрочем, возможно, что гипотеза о древнем мире РНК как предшественнике современного живого мира так и не сможет получить достаточного обоснования для преодоления основной трудности - научно правдоподобного описания механизма перехода от РНК и ее репликации к биосинтезу белка. Имеется привлекательная и детально продуманная альтернативная гипотеза А.Д. Альтштейна (Институт биологии гена РАН), в которой постулируется, что репликация генетического материала и его трансляция - синтез белка - возникали и эволюционировали одновременно и сопряженно, начиная с взаимодействия абиогенно синтезирующихся олигонуклеотидов и аминоацил-нуклеотидилатов - смешанных ангидридов аминокислот и нуклеотидов . Но это уже следующая сказка... ("И Шахразаду застигло утро, и она прекратила дозволенные речи" .)
Литература
1. Watson J.D., Crick F.H.C.
Molecular structure of
nucleic acids // Nature. 1953. V. 171. P. 738-740. 2. Watson J.D., Crick F.H.C.
Genetic implications of
the structure of deoxyribose nucleic acid // Nature 1953 V. 171. P. 964-967. 3. Спирин А.С.
Современная биология и биологическая
безопасность // Вестник РАН. 1997. № 7. 4. Spirin A.S.
On macromolecular structure of native
high-polymer ribonucleic acid in solution // Journal of Molecular Biology.
1960. V. 2. P. 436-446. 5. Kirn S.H., Suddath F.L., Quigley GJ. et al.
Three-dimensional tertiary structure of yeast phenylalanine transfer RNA //
Science. 1974. V. 185. P. 435-40. 6. Robertas J.D., Ladner J.E., Finch J.T. et al.
Structure of yeast phenylalanine tRNA at 3 A resolution // Nature. 1974.
V. 250. P. 546-551. 7. Vasiliev V.D., Serdyuk I.N., Gudkov A.T., SPIRin
A.S.
Self-organization of ribosomal RNA // Sturcture, Function and
Genetics of Ribosomes / Eds. Hardesty B. and Kramer G. New York:
Springer-Verlag, 1986. P. 129-142. 8. Baserga SJ., Steitz J.A.
The diverse world of small
ribo-nucleoproteins // The RNA World / Eds. Gesteland R.F. and Atkins J.F. New
York: Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1993. P.
359-381. 9. Kruger К., Grabowski PJ., Zaug AJ. et al.
Self-splicing RNA: Autoexcision and autocyclization of the ribosomal RNA
intervening sequence of Tetrahymena
// Cell. 1982. V. 31. P. 147-157. 10. Guerrier-Takada С., Gardiner К., Marsh Т. et al.
The RNA moiety of ribonucleases P is the catalytic subunit of the enzyme
// Cell. 1983. V. 35. P. 849-857. 11. Опарин А.И.
Происхождение жизни. М.: Московский
рабочий, 1924. 12. Опарин А.И.
Возникновение жизни на Земле (3-е
изд.). М.: Изд-во АН СССР, 1957. 13. Woese С.
The evolution of the genetic code // The
Genetic Code. New York: Harper & Row, 1967. P. 179-195. 14. Crick F.H.C.
The origin of the genetic code //
Journal of Molecular Biology. 1968. V. 38. P. 367-379. 15. Orgel L.E.
Evolution of the genetic apparatus //
Journal of Molecular Biology. 1968. V. 38. P. 381-393. 16. Gilbert W.
The RNA world // Nature. 1986. V 319
P. 618. 17. Joyce G.F., Orgel L.E.
Prospects for
understanding the origin of the RNA world // The RNA World / Eds. Gesteland
R.F. and Atkins J.F.
New York: Cold Spring Harbor Laboratory Press, Cold
Spring Harbor, 1993 P 1-25. 18. Chetverina H.V., Demidenko A.A., Ugarov V.I.,
Chetverin A.B.
Spontaneous rearrangements in RNA sequences // FEBS
Letters. 1999. V. 450. P. 89-94. 19. Bartel D.P., Szostak J.W.
Isolation of new
ribozymes from a large pool of random sequences // Science. 1993. V. 261. P.
1411-1418. 20. Ekland E.H., Bartel D.P.
RNA-catalysed RNA
polymerization using nucleoside triphosphates // Nature. 1996 V. 382. P.
373-376. 21. Orgel L.E.
The origin of life - a review of facts
and speculations //Trends in Biochemical Sciences. 1998. V. 23. p. 491-495. 22. Альтштейн А.Д.
Происхождение генетической
системы: гипотеза прогенов // Молекулярная биология. 1987. Т. 21. С. 309-322.
Общепринятого определения жизни нет. Нам известна только одна жизнь - земная, и мы не знаем, какие из ее свойств являются обязательными для любой жизни вообще. Можно предположить два таких свойства. Это, во-первых, наличие наследственной информации, во-вторых - активное осуществление функций, направленных на самоподдержание и размножение, а также на получение энергии, необходимой для выполнения всей этой работы.
Все живое на Земле справляется с перечисленными задачами при помощи трех классов сложных органических соединений: ДНК, РНК и белков. ДНК взяла на себя первую задачу - хранение наследственной информации. Белки отвечают за вторую: они выполняют все виды активных «работ». Разделение труда у них очень строгое.
Молекулы третьего класса веществ - РНК - служат посредниками между ДНК и белками, обеспечивая считывание наследственной информации. При помощи РНК осуществляется синтез белков в соответствии с записанными в молекуле ДНК «инструкциями». Некоторые из функций, выполняемых РНК, очень похожи на функции белков (активная работа по прочтению генетического кода и синтезу белка), другие напоминают функции ДНК (хранение и передача информации). И все это РНК делает не в одиночку, а при активном содействии со стороны белков. На первый взгляд РНК кажется «третьей лишней». В принципе нетрудно представить себе организм, в котором РНК вовсе нет, а все ее функции поделили между собой ДНК и белки. Правда, таких организмов в природе не существует.
Какая из трех молекул появилась первой? Одни ученые говорили: конечно, белки, ведь они выполняют всю работу в живой клетке, без них жизнь невозможна. Им возражали: белки не могут хранить наследственную информацию, а без этого жизнь и подавно невозможна! Значит, первой была ДНК!
Ситуация казалась неразрешимой: ДНК ни на что не годна без белков, белки - без ДНК. Получалось, что они должны были появиться вместе, одновременно, а это трудно себе представить. Про «лишнюю» РНК в этих спорах почти забыли.
Потом, правда, выяснилось, что у многих вирусов наследственная информация хранится в виде молекул РНК, а не ДНК. Но это посчитали курьезом, исключением. Переворот произошел в 80-х годах XX века, когда были открыты рибозимы - молекулы РНК с каталитическими свойствами. Рибозимы - это РНК, выполняющие активную работу, то есть то, что должны делать белки.
В итоге РНК из «почти лишней» стала «почти главной». Оказалось, что она, и только она, может выполнять сразу обе главные жизненные задачи - и хранение информации, и активную работу. Стало ясно, что возможен полноценный живой организм, не имеющий ни белков, ни ДНК, в котором все функции выполняются только молекулами РНК. Конечно, ДНК лучше справляется с задачей хранения информации, а белки - с «работой», но это уже детали. РНК-организмы могли приобрести белки и ДНК позже, а поначалу обходиться без них.
Так появилась теория РНК-мира, согласно которой первые живые существа были РНК-организмами без белков и ДНК. А первым прообразом будущего РНК-организма мог стать автокаталитический цикл, образованный самовоспроизводящимися молекулами РНК - рибозимами, способными катализировать синтез собственных копий.
Лично я считаю теорию РНК-мира одним из самых выдающихся достижений теоретической мысли в биологии. По правде сказать, могли бы до этого додуматься и раньше. Ведь два вида рибозимов были известны еще с 60-х годов ХХ века, хотя их не называли тогда рибозимами. Это рибосомные РНК (рРНК), из которых сделаны молекулярные «машинки» для трансляции (синтеза белка) - рибосомы, и транспортные РНК (тРНК), которые подносят нужные аминокислоты к рибосомам в ходе трансляции.
Теория РНК-мира, вначале чисто умозрительная, быстро «обрастает» экспериментальными данными. Химики научились получать рибозимы чуть ли не с любыми желаемыми характеристиками. Делается это так. Например, мы хотим создать молекулу РНК, которая способна безошибочно узнавать вещество Х и связываться с ним. Для этого синтезируют большое количество разных цепочек РНК, соединяя рибонуклеотиды друг с другом в случайном порядке. Раствор, содержащий полученную смесь молекул РНК, наливают на поверхность, покрытую веществом Х. После этого остается лишь отобрать и исследовать те молекулы РНК, которые прилипли к поверхности. Технология незамысловата, но она действительно работает. Примерно таким способом получены рибозимы, катализирующие синтез нуклеотидов, присоединяющие аминокислоты к РНК и выполняющие множество других биохимических функций.
Среди современных концепций зарождения жизни одно из доминирующих положений занимает теория РНК-мира . Попробуем разобраться, что же это такое.
Открытия в молекулярной биологии прошлого столетия привели человечество к пониманию устройства жизни на химическом уровне. Выяснилось, что основу жизнедеятельности любого организма составляют две группы веществ-биополимеров: белки и нуклеиновые кислоты.
Белки, чьи длинные, хитроумно свернутые цепи состоят из десятков и сотен последовательно связанных аминокислот, выполняют в клетке роль рабочих инструментов и универсального строительного материала. Белки-ферменты ускоряют и направляют все химические реакции, протекающие в клетке, формируя ее облик.
Но белки - временные инструменты, потребность в которых постоянно изменяется по ходу жизни организма. Для хранения же информации о белках, а значит, и о строении самого организма природа использует нуклеиновые кислоты - ДНК (дезоксирибонуклеиновую кислоту) и РНК (рибонуклеиновую кислоту). Эти длинные молекулы, построенные из сцепленных между собой четырех видов нуклеотидов, очень похожи по строению, но обладают разными свойствами. Две направленные в разные стороны цепи ДНК формируют жесткую и стабильную двойную спираль длиной в миллионы пар нуклеотидов. РНК же образует сравнительно короткие цепи, подверженные разнообразным химическим реакциям и заплетенные петлями сами на себя.
Структура молекулы ДНК. Изображение: Richard Wheeler / Wikimedia
Столь различная структура объяснила ученым принципиально разные функции ДНК и РНК. ДНК оказалась надежным, долговременным хранилищем информации о белках организма, а РНК - мобильным, коротко живущим переносчиком информации. Она синтезируется белками-полимеразами по ДНК-матрице и отвечает за расшифровку информации, записанной в ДНК, а также за сборку белков по ДНК-чертежу.
Весь этот ворох знаний был накоплен учеными к середине 60-х годов прошлого столетия, став предтечей настоящей биотехнологической революции. Но одновременно он поставил ученых, мучающихся над проблемой зарождения жизни, перед парадоксом.
Для существования первых «живых», то есть способных к размножению и самоподдержанию биохимических систем, достаточно ДНК, РНК и белка. С ролью РНК все вроде бы понятно - типичная молекула на побегушках, которая ничего толком не умеет и не решает, но необходима для переноса информации из ДНК и работы механизмов сборки белка. А вот белки и ДНК явно должны были занимать центральное место в картине доисторического мира.
Информация о структуре белков-катализаторов, умеющих все на свете, способна сохраняться, только будучи записанной в структуре ДНК. Одновременно стабильная ДНК, отлично сохраняя информацию, не способна на самостоятельные химические превращения, кроме, разве что, медленного распада. Что же появилось в эволюции раньше - умелые, короткоживущие белки или надежная, но беспомощная ДНК? Одно никак не может появиться без другого, а случайное одномоментное зарождение сложной ДНК-РНК-белковой самовоспроизводящейся системы казалось невероятным.
Тут взгляды ученых и обратились на РНК. РНК не стабильна и ужасно плохо хранит информацию, но все же хранит ее. А что если допустить, что заплетенные в витиеватые петли цепи РНК смогут работать наподобие белков-ферментов, катализируя, то есть ускоряя, биохимические реакции? Пусть они бы справлялись с этой задачей в сотни раз хуже белков, но гипотетически такие РНК-катализаторы могли бы устойчиво существовать и размножаться на поверхности древней Земли еще до появления белков и ДНК. А их химическая нестабильность была бы даже плюсом, приводя к бешеному темпу эволюции первобытной РНК-фауны.

Структура молекулы предшественника матричной РНК. Изображение: Vossman / Wikimedia
Смелая гипотеза оказалась пророческой, в начале 80-х были найдены первые рибозимы - биокатализаторы на основе РНК. Чуть позже ученые получили аптамеры - молекулы РНК, способные избирательно связывать определенные вещества . Оказалось, что РНК может выполнять работу как по биокатализу, так и по молекулярному распознаванию. Да, у нее это получается хуже, чем у белков, но все же получается.
С тех пор ученые не оставляют настойчивых попыток получить в лаборатории рибозим, способный к устойчивому копированию (репликации) молекул РНК любой структуры. Появившись на заре эволюции, аналогичный рибозим стал бы настоящим «ядром» гипотетического РНК-мира, а его получение было бы осязаемым подтверждением пока еще умозрительной гипотезы.
За годы исследований были получены рибозимы-лигазы, способные сшивать молекулы РНК между собой, и даже рибозимы-полимеразы, копирующие небольшие, однородные по своему нуклеотидному составу фрагменты РНК. Но на всех сложных, способных к биокатализу и молекулярному распознаванию последовательностях они упрямо буксовали, отказываясь работать.
И вот недавно в авторитетном журнале PNAS была опубликована статья о получении первого рибозима, уверенно копирующего РНК-матрицы любого нуклеотидного состава. В ходе экспериментов ученые подменили собой эволюцию: путем искусственного отбора в пробирке им удалось создать рибозим, копирующий РНК с недоступной ранее точностью.
Каждый из 24 раундов мутации-отбора начинался с копирования уже существующего фермента в биохимическом процессе, получившем название рибоПЦР. Эта реакция - аналог хорошо известной полимеразной цепной реакции (ПЦР), позволяющей за несколько часов синтезировать миллионы копий нужного фрагмента ДНК. Для того чтобы в системе появился материал для искусственного отбора, реакция была модифицирована в сторону уменьшения точности копирования. Частота ошибок достигала 10% в пересчете на отдельный нуклеотид. Благодаря этому запланированному случайному мутагенезу ученым удалось получить 10 14 (100 триллионов!) различных вариантов исходного рибозима. После завершения реакции мутантные рибозимы придирчиво отбирались учеными: в следующий раунд мутации проходили только самые быстрые и точные рибозимы, способные к наилучшему копированию матрицы.
После завершения этой кропотливой работы исследователи получили рибозим, названный 24-3 полимераза. Впервые в руки ученых попал рибозим, способный реплицировать небольшие цепи РНК любой последовательности. С его помощью удалось реплицировать несколько аптамеров. Затем неутомимой полимеразой был копирован каталитически активный рибозим-лигаза. Но настоящим достижением стало то, что с помощью 24-3 полимеразы удалось реплицировать одну из транспортных РНК. Эти крупные, хитро заплетенные в фигуру наподобие клеверного листа молекулы РНК переносят звенья-аминокислоты к месту сборки белковых цепей и являются важнейшим компонентом аппарата синтеза белка.
Скорость работы полученного рибозима оказалась крайне мала, а производительность несравнима с природными белками-полимеразами, но главное - он был получен, и он работает. Теперь для доказательства возможности существования древнего РНК-мира ученым остался последний шаг - создать рибозим, способный устойчиво реплицировать сам себя. Сделав его, человечество получит в пробирке колонию самокопирующихся молекул РНК - потенциальный аналог первой формы жизни на нашей планете.
Несколько месяцев работы позволили исследователям вплотную приблизиться к созданию искусственного прототипа первобытной жизни. Что же могло получится у естественного отбора за сотни миллионов лет? Еще никогда мы не были так близки к ответу на этот вопрос.
Текущая версия страницы пока не проверялась
Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от, проверенной 15 июля 2019; проверки требуют.
Мир РНК - гипотетический этап возникновения жизни на Земле , когда как функцию хранения генетической информации, так и катализ химических реакций выполняли ансамбли молекул рибонуклеиновых кислот . Впоследствии из их ассоциаций возникла современная ДНК -РНК -белковая жизнь, обособленная мембраной от внешней среды. Идея мира РНК была впервые высказана Карлом Вёзе в 1968 году , позже развита Лесли Орджелом и окончательно сформулирована Уолтером Гильбертом в 1986 году .
В живых организмах практически все процессы происходят в основном благодаря ферментам белковой природы. Белки, однако, не могут самореплицироваться и синтезируются в клетке de novo на основании информации, заложенной в ДНК . Но и удвоение ДНК происходит только благодаря участию белков и РНК. Образуется замкнутый круг, из-за которого в рамках теории самозарождения жизни приходилось признать необходимость не только абиогенного синтеза обоих классов молекул, но и спонтанного возникновения сложной системы их взаимосвязи.
Таким образом, РНК могли существовать полностью автономно, катализируя «метаболические» реакции, например, синтеза новых рибонуклеотидов и самовоспроизводясь, сохраняя из «поколения» в «поколение» каталитические свойства. Накопление случайных мутаций привело к появлению РНК, катализирующих синтез определённых белков, являющихся более эффективным катализатором, в связи с чем эти мутации закреплялись в ходе естественного отбора. С другой стороны возникли специализированные хранилища генетической информации - ДНК. РНК сохранилась между ними как посредник.
Следы мира РНК остались в современных живых клетках , причём РНК участвует в критически важных процессах жизнедеятельности клетки:
В 2017 году была показана возможность самосборки уридина из рибозы, урацила и фосфорной кислоты.
Способность молекул РНК к эволюции была наглядно продемонстрирована в ряде экспериментов. Ещё до открытия каталитической активности РНК такие эксперименты проводили Лесли Орджел с коллегами в Калифорнии. Они добавляли к пробирке с РНК яд - бромид этидия , ингибирующий синтез РНК. Сначала темп синтеза был замедлен ядом, но примерно после девяти «пробирочных поколений» эволюции в процессе естественного отбора вывелась новая порода РНК, стойкая к яду. Путём последовательного удвоения доз яда была выведена порода РНК, стойкая к очень высоким его концентрациям. Всего в эксперименте сменилось 100 пробирочных поколений (и намного больше поколений РНК, так как поколения сменялись и внутри каждой пробирки). Хотя в этом эксперименте РНК-репликаза добавлялась в раствор самими экспериментаторами, Орджел обнаружил, что РНК способны и к спонтанному самокопированию, без добавления фермента, правда, намного медленнее.
Дополнительный эксперимент был позже проведён в лаборатории немецкой школы Манфреда Ейгена. Он обнаружил спонтанное самозарождение молекулы РНК в пробирке с субстратом и РНК-репликазой. Она была создана постепенно нарастающей эволюцией.
После открытия каталитической активности РНК (рибозимов) их эволюция в автоматизированном устройстве под управлением компьютера наблюдалась в экспериментах Брайана Пегеля и Джеральда Джойса из Исследовательского института имени Скриппса в Калифорнии в 2008 году. Фактором, играющим роль давления отбора, являлась ограниченность субстрата, куда входили олигонуклеотиды, которые рибозим распознавал и присоединял к себе, и нуклеотиды для синтеза РНК и ДНК. При построении копий иногда случались дефекты - мутации - влияющие на их каталитическую активность (для ускорения процесса несколько раз смесь подвергалась мутированию с помощью полимеразной цепной реакции с использованием «неточных» полимераз). По этому признаку и происходил отбор молекул: наиболее быстро копирующиеся молекулы быстро начинали доминировать в среде. Затем 90 % смеси удалялось, а вместо этого добавлялась свежая смесь с субстратом и ферментами, и цикл повторялся снова. За 3 суток каталитическая активность молекул за счёт всего 11 мутаций увеличилась в 90 раз.
Эти эксперименты доказывают, что первым молекулам РНК не нужно было обладать достаточно хорошими каталитическими свойствами. Они развились потом в ходе эволюции под действием естественного отбора.
В 2009 году канадские биохимики из Монреальского университета К. Боков и С. Штейнберг, изучив основную составляющую рибосомы бактерии Escherichia coli , молекулу 23S-рРНК, показали, каким образом из относительно небольших и простых рибозимов мог развиться механизм белкового синтеза . Молекула была подразделена на 60 относительно самостоятельных структурных блоков, основным из которых является каталитический центр (пептидил-трансферазный центр, PTC, peptidyl-transferase centre), ответственный за транспептидацию (образование пептидной связи). Было показано, что все эти блоки можно последовательно отсоединять от молекулы без разрушения её оставшейся части до тех пор, пока не останется один лишь транспептидационный центр. При этом он сохраняет способность катализировать транспептидацию. Если каждую связь между блоками молекулы представить в виде стрелки, направленной от того блока, который при отрыве не разрушается, к тому блоку, который разрушается, то такие стрелки не образуют ни одного замкнутого кольца. Если бы направление связей было случайным, вероятность этого составляла бы менее одной миллиардной. Следовательно, такой характер связей отражает последовательность постепенного добавления блоков в процессе эволюции молекулы, который исследователям удалось детально реконструировать. Таким образом, у истоков жизни мог стоять сравнительно простой рибозим - PTC-центр молекулы 23S-рРНК, к которому затем добавлялись новые блоки, совершенствуя процесс синтеза белка. Сам PTC состоит из двух симметричных лопастей, каждая из которых удерживает CCA"-хвостик одной молекулы тРНК . Предполагается, что такая структура возникла в результате дупликации (удвоения) одной исходной лопасти. Методом искусственной эволюции были получены функциональные РНК (рибозимы), способные катализировать транспептидацию. Структура этих искусственно выведенных рибозимов очень близка к структуре той проторибосомы, которую «вычислили» авторы.
О том, как выглядели самовоспроизводящиеся РНК системы, есть разные предположения. Чаще всего постулируется необходимость агрегирующих РНК мембран или размещения РНК на поверхности минералов и в поровом пространстве рыхлых пород. В 1990-е годы А. Б. Четвериным с сотрудниками была показана способность РНК формировать молекулярные колонии на гелях и твёрдых субстратах при создании им условий для репликации. Происходил свободный обмен молекулами, которые при столкновении могли обмениваться участками, что показано экспериментально. Вся совокупность колоний в связи с этим быстро эволюционировала .
После возникновения белкового синтеза колонии, умеющие создавать ферменты, развивались успешнее. Ещё более успешными стали колонии, сформировавшие более надёжный механизм хранения информации в ДНК и, наконец, отделившиеся от внешнего мира липидной мембраной, препятствующей рассеиванию своих молекул.
Пре-биотические модели, при которых создаются нуклеотиды , несовместимы с условиями, необходимыми для создания сахаров (из за большой концентрации формальдегида). Так что они должны быть синтезированы в разных местах, а затем перенесены в какое-то одно место. Однако, они не реагируют в воде. Безводные реакции легко связывают пурины с сахарами, но только 8 % из них соединяют правильный углеродный атом на сахаре с правильным атомом азота на основе. Пиримидины , однако, не будут реагировать с рибозой, даже в безводных условиях.
Кроме того, необходимые для синтеза фосфаты в природе встречаются крайне редко, так как они легко выпадают в осадок. При введении фосфата, последний должен быстро соединяться с правильной гидроксильной группой нуклеотида.
Чтобы нуклеотиды могли сформировать РНК, они сами должны быть активизированы. Активизированыe пуриновые нуклеотиды формируют маленькие цепочки на существующем шаблоне пиримидиновых РНК, но этот процесс не идет наоборот потому что пиримидиновые нуклеотиды не полимеризуются так легко.
Другой гипотезой абиогенного синтеза РНК, призванной решить проблему низкой оценочной вероятности синтеза РНК, является гипотеза мира полиароматических углеводородов , предложенная в 2004 году и предполагающая синтез молекул РНК на основе стека из полиароматических колец.
Фактически, обе гипотезы «пре-РНК миров» не отвергают гипотезу мира РНК, а модифицируют её, постулируя первоначальный синтез реплицирующихся макромолекул РНК в первичных метаболических компартментах, либо на поверхности ассоциатов, отодвигая «мир РНК» на вторую стадию
Жизнь начиналась с РНК
Исследования нуклеиновых кислот являются одной из самых «горячих точек» в биологии. Благодаря уникальным свойствам РНК находят все более широкое применение в медицине и технике. Но знает об этом пока лишь узкий круг специалистов.
Рибонуклеиновой кислоте, иначе - РНК - не повезло. Она не пользуется такой широкой известностью, как ее близкий «родственник» - ДНК, несмотря на большое химическое сходство. Однако открытия последних двадцати лет радикально поменяли наши взгляды на роль и функции этих, как выяснилось, очень «умелых» молекул. Плодом этих открытий стала принципиально новая идея о том, что современной жизни предшествовал совершенно самодостаточный древний «мир РНК».
Как это обычно бывает, новое знание, расширяя горизонт, породило и массу новых вопросов. Каковы были механизмы «эволюции» в мире РНК? Зачем, откуда и как появились ДНК и белки? Как произошел переход от «мира РНК» к современному миру? О поисках, которые ведутся в этом направлении, читателям рассказывают академик Валентин Викторович Власов и его сын, кандидат химических наук, Александр Власов.
Почему в цикле статей, посвященных проблеме возникновения жизни, появляется статья об РНК, а не о других, более известных органических молекулах - ДНК или белках? Возможно, наши читатели слышали и об РНК, но вот что? Уверены, ничего примечательного - по одной простой причине: пока лишь специалисты-биологи знают, что именно РНК являются «волшебными» молекулами, давшими начало жизни. Что когда-то в древности, на только что остывшей Земле, возник и существовал загадочный «мир РНК»…Прежде чем отправиться к «началу начал», давайте запасемся необходимыми знаниями о строении нуклеи-новых кислот - ДНК (дезоксирибонуклеиновой) и РНК (рибонуклеиновой). По своему химическому составу РНК является двойняшкой, хотя и не полным близнецом, ДНК, основного хранителя генетической информации в живой клетке. Нуклеиновые кислоты представляют собой полимерные макромолекулы, состоящие из отдельных звеньев - нуклеотидов . Скелетом макромолекулы являются молекулы пятиуглеродного сахара, соединенные остатками фосфорной кислоты. К каждой молекуле сахара присоединяется одно азотистое основание. Нуклеотиды, которые различаются между собою только разными азотистыми основаниями, обозначаются буквами A, U, G, C (в РНК) и A, T, G, C (в ДНК).
Честно говоря, насчет РНК никто не задумывался долгие годы. Существовала догма, что вот есть клетка, есть хромосомы, в которых есть ДНК - хранитель генетической информации.
В конце концов, на рибосомах синтезируются белки. А РНК - она где-то в промежутке, переносчик информации от ДНК - и только. А потом посыпались открытия, которые заставили совершенно по-другому взглянуть на РНК
Главное отличие нуклеиновых кислот заключается в их углеводной компоненте. В РНК сахар - рибоза, а в ДНК - дезоксирибоза: там, где у ДНК имеется атом водорода (Н), у РНК стоит оксигруппа (ОН). Результаты таких незначительных, на неискушенный взгляд, различий поражают. Так, ДНК существуют в основном в форме всем известных жестких спиралей, в которых две цепи ДНК удерживаются вместе за счет образования водородных связей между комплементарными нуклеотидами.
РНК также могут формировать спирали из двух цепочек, похожие на спирали ДНК, однако в большинстве случаев РНК существуют в виде сложных структур-клубков. Структуры эти формируются не только за счет образования упомянутых водородных связей между разными участками РНК, но и благодаря оксигруппе рибозы, которая может образовывать дополнительные водородные связи и взаимодействовать с фосфорной кислотой и ионами металлов. Глобулярные структуры РНК не только внешне напоминают белковые структуры, но и приближаются к ним по свойствам: они могут взаимодействовать с самыми разными молекулами, как маленькими, так и полимерными.
Кого Считать «Живым»?
Почему же именно РНК мы называем праматерью ныне существующей жизни? Чтобы ответить на этот вопрос, давайте разберемся, где проходит граница между живым и неживым.Поскольку над проблемой происхождения жизни работают ученые из разных областей, каждый оперирует терминами близкой ему науки. Химики обязательно вспомнят слово «катализатор», математики - «информация». Биологи будут считать живой систему, содержащую вещество (генетическую программу), которое может копироваться (или, по-простому, размножаться). При этом необходимо, чтобы в ходе такого копирования могли происходить некоторые изменения наследственной информации и возникать новые варианты систем, т. е. должна существовать возможность эволюции . Еще биологи обязательно заметят, что такие системы должны быть пространственно обособлены. Иначе возникшие более прогрессивные системы не смогут воспользоваться своими преимуществами, поскольку их более эффективные катализаторы и другие продукты будут беспрепятственно «уплывать» в окружающую среду.
Каким же образом первые молекулярные системы были обособлены от окружающей среды? Колонии молекул могли, например, удерживаться вместе за счет адсорбции на какой-нибудь минеральной поверхности или пылевых частицах. Однако возможно, что уже самые примитивные системы располагали, подобно современным живым клеткам, настоящей мембранной оболочкой. Дело в том, что такая «протоклетка» с липидной мембраной может образоваться очень просто. Многие молекулы с заряженными группами (например, жирные кислоты) в водной среде образуют микроскопические пузырьки - липосомы . Это слово должно быть хорошо известно прекрасной половине наших читателей: липосомы широко используются в косметических кремах - крохотные жировые капсулы начиняются витаминами и другими биологически активными веществами. А вот чем были наполнены древние «протоклетки»? Оказалось, что на роль «начинки» претендуют именно РНК.
РНК умеет все?
Жизнь, без сомнения, должна была начаться с образования «умелых» молекул, которые могли бы сами себя размножать и выполнять все другие «хозяйственные работы», необходимые для существования клетки. Однако на роль таких умельцев не подходит ни ДНК, ни белок. ДНК - отличный хранитель генетической информации, но сама себя размножать не умеет. Белки - непревзойденные катализаторы, но не могут работать в качестве «генетических программ». Возникает парадокс курицы и яйца: ДНК не может образоваться без белка, а белок - без ДНК. И только РНК, как выяснилось, может ВСЕ. Но не будем забегать вперед.Рассмотрим давно известные функции РНК, связанные с работой (экспрессией ) гена в клетке. При включении гена сначала происходит локальное расплетение ДНК и синтезируется РНК-копия генетической программы. В результате сложных обработок ее специальными белками получается матричная РНК (мРНК ), которая и явля-ется программой для синтеза белка. Эта РНК переносится из ядра в цитоплазму клетки, где она связывается со специальными клеточными структурами - рибосомами , настоящими молекулярными «машинами» для синтеза белка. Белок синтезируется из активированных аминокислот, присоединенных к особым транспортным РНК (тРНК), причем каждая из аминокислот присоединена к своей специфической тРНК. Благодаря тРНК аминокислота фиксируется в каталитическом центре рибосомы, где она «пришивается» к синтезируемой белковой цепи. Из рассмотренной последовательности событий видно, что молекулы РНК играют ключевую роль в декодировании генетической информации и биосинтезе белка.
Чем больше углублялись в изучение различных биосинтетических процессов, тем чаще обнаруживали ранее неизвестные функции РНК. Оказалось, что кроме процесса транскрипции (синтеза РНК путем копирования участка ДНК) в ряде случаев, наоборот, может происходить синтез ДНК на РНК-матрицах. Этот процесс, названный обратной транскрипцией , используют в ходе своего развития многие вирусы, в том числе печально известные онкогенные вирусы и ВИЧ-1, вызывающий СПИД.
Таким образом, выяснилось, что поток генетической информации не является, как первоначально считалось, однонаправленным - от ДНК к РНК. Роль ДНК как изначально главного носителя генетической информации стала подвергаться сомнению. Тем более что многие вирусы (гриппа, клещевого энцефалита и другие) вообще не используют ДНК в качестве генетического материала, их геном построен исключительно из РНК. А далее посыпались одно за другим открытия, которые заставили совершенно по-другому взглянуть на РНК.
На Все «Молекулы» Мастер
Наиболее удивительным было открытие каталитической способности РНК. Прежде считалось, что катализировать реакции умеют только белки, ферменты. Ученые, например, никак не могли выделить ферменты, осуществляющие разрезание и сшивание некоторых РНК. После длительных исследований выяснилось, что РНК прекрасно справляются с этим сами. Структуры РНК, действующие подобно ферментам, назвали рибозимами (по аналогии с энзимами , белками-катализаторами). Вскоре было обнаружено множество разнообразных рибозимов. Особенно широко их используют для манипулирования своими РНК вирусы и другие простые инфекционные агенты. Таким образом, РНК оказались мастерами на все руки: они могут выступать в роли носителей наследственной информации, могут служить катализаторами, транспортными средствами для аминокислот, образовывать высокоспецифичные комплексы с белками.Окончательная уверенность в том, что «мир РНК» действительно существовал, наступила после выявления деталей строения кристаллов рибосом методом рентгеноструктурного анализа. Ученые рассчитывали обнаружить там белок, катализирующий сшивание аминокислот в белковую последовательность. Каково же было их удивление, когда выяснилось, что в каталитическом центре рибосом белковых структур нет совсем, что он полностью построен из РНК! Оказалось, что все ключевые стадии биосинтеза белка осуществляются молекулами РНК. Точка в дискуссии о возможности существования «мира РНК» как особой стадии биологической эволюции была поставлена.
Конечно, полную картину еще предстоит реконструировать - осталось много нерешенных вопросов. Например, в современной клетке активацию аминокислот и их присоединение к соответствующим тРНК осуществляют специфичные белки-ферменты. Возникают вопросы: могла ли эта реакция осуществляться без участия белков, только с помощью РНК? Могли ли сами РНК катализировать синтез РНК из нуклеотидов или присоединение азотистых оснований к сахару? В общем-то, после открытия рибозимов такие потенциальные способности РНК уже не вызывали особых сомнений. Но наука требует, чтобы гипотезы экспериментально подтверждались.
Дарвиновская Эволюция в Пробирке
Хороший метод зачастую позволяет осуществить революцию в науке. Именно так можно сказать о методе полимеразной цепной реакции (ПЦР) , который позволяет размножать нуклеиновые кислоты в неограниченных количествах. Кратко опишем суть метода. Для размножения ДНК в методе ПЦР используются ферменты ДНК- полимеразы , т. е. те самые ферменты, которые при размножении клеток синтезируют из активированных мономеров-нуклеотидов комплементарные цепочки ДНК.При методе ПЦР в пробирку с ДНК вносят смесь активированных нуклеотидов, фермент ДНК-полимеразу и так называемые праймеры - олигонуклеотиды, комплементарные концам размножаемой ДНК. При нагревании раствора цепи ДНК расходятся. Затем, при охлаждении, с ними связываются праймеры, образуя короткие фрагменты спиральных структур. Фермент присоединяет к праймерам нуклеотиды и собирает цепочку, комплементарную цепочке исходной ДНК. В результате реакции из одной двуцепочечной ДНК получается две. Если повторить процесс, получится четыре цепочки, а после n повторений - 2 n молекул ДНК. Все очень просто.
Изобретение ПЦР и разработка методов химического синтеза ДНК позволили создать потрясающую технологию молекулярной селекции. Принцип молекулярной селекции тоже прост: сначала синтезируется множество молекул, обладающих разными свойствами (так называемая молекулярная библиотека ), а затем из этой смеси отбираются молекулы с желаемым свойством.
Библиотеки нуклеиновых кислот - это смеси молекул, имеющих одинаковую длину, но отличающихся последовательностью нуклеотидов. Получить их можно в том случае, если при химическом синтезе на авто-матическом синтезаторе добавлять на каждой стадии удлинения нуклеотидной последовательности одно-временно все четыре нуклеотида. Каждый из них будет включаться в растущую нуклеиновую кислоту с равной вероятностью, в результате чего на каждом этапе присоединения будет получаться 4 варианта последовательностей. Если таким образом синтезировать нуклеиновую кислоту длиной в n звеньев, то разнообразие полученных молекул составит 4 в степени n. Поскольку обычно используются участки длиной 30-60 мономеров, то в результате синтеза получается от 4 30 до 4 60 разных молекул! Цифры, привычные разве что для астрономов.
Так как в зависимости от состава нуклеиновые кислоты сворачиваются в разные пространственные структуры, синтез статистических последовательностей дает огромное множество молекул, различающихся по свойствам. С образовавшихся ДНК - с помощью фермента РНК-полимеразы - считывается РНК. В результате получается библиотека уже одноцепочечных РНК. Далее производится процедура отбора: раствор РНК пропускается через колонку, в которой находится нерастворимый носитель с химически присоединенными молекулами-мишенями, чтобы «выловить» так называемый будущий аптамер , т. е. РНК, способную связывать определенные молекулы. Затем колонку промывают для удаления несвязавшихся РНК, а затем смывают РНК, задержавшиеся на колонке за счет связывания с целевыми молекулами (это можно сделать, например, нагревая колонку).
С выделенных РНК с помощью обратной транскрипции делают ДНК-копии и получают из них обычные двуцепочечные молекулы ДНК. С последних же можно считывать искомые РНК-аптамеры, а затем - размножать их методом ПЦР в неограниченных количествах. Конечно, так происходит в идеальном случае, на практике все получается сложнее. Обычно исходный препарат РНК содержит огромный избыток «по-сторонних» молекул, избавиться от которого трудно. Поэтому полученную РНК вновь и вновь пропускают через колонку, чтобы выделить РНК, образующие самые прочные комплексы с целевыми молекулами.
С помощью такого метода были получены тысячи разных РНК-аптамеров, которые образуют специфические комплексы с различными органическими соединениями и молекулами.
Рассмотренная схема молекулярной селекции может быть применена для получения молекул с любыми свойствами. Например, были получены РНК, способные катализировать реакции синтеза РНК и белков: присоединение азотистых оснований к рибозе, полимеризацию активированных нуклеотидов на цепочках РНК, присоединение аминокислот к РНК. Эти исследования еще раз подтвердили, что в условиях предбиологической эволюции из случайных полимеров могли возникать молекулы РНК
со специфическими структурами и функциями.
Делайте Ваш Заказ!
Метод молекулярной селекции обладает очень большими возможностями. С его помощью можно решать задачи поиска нужных молекул даже в том случае, если исходно нет идеи, как такие молекулы должны быть устроены. Однако, если придумать процедуру отбора, их можно выделить по принципу требуемых свойств, а затем уже заняться и вопросом, как эти свойства достигаются. Продемонстрируем это на примере выделения РНК, способных связываться с клеточными мембранами и модулировать их проницаемость.Древние рибоциты должны были поглощать «питательные» вещества из окружающей среды, удалять продукты метаболизма и делиться в ходе размножения.
И все эти процессы требуют управления проницаемостью мембран. Поскольку мы полагаем, что никаких других функциональных молекул, кроме РНК, в рибоцитах не было, какие-то РНК обязательно должны были взаимодействовать с мембранами. Однако с химической точки зрения они совершенно не подходят для роли регуляторов проницаемости мембран.
Мембраны современных клеток и липосом, построенные из жирных кислот, несут отрицательный заряд. Поскольку РНК также заряжены отрицательно, то по закону Кулона они должны отталкиваться от липидной поверхности и тем более не могут проникать в глубь липидного слоя. Единственный известный способ взаимодействия нуклеиновых кислот с поверхностью мембран - через двухзарядные ионы металлов. Эти положительно заряженные ионы могут играть роль мостиков, располагаясь между отрицательно заряженными группами на поверхности мембраны и фосфатными группами нуклеиновой кислоты. Поскольку такие мостиковые взаимодействия достаточно слабые, с мембраной может связаться только очень большая нуклеиновая кислота благодаря множеству слабых связей с поверхностью мембраны. Так маленькие враги привязали Гулливера к земле множеством тоненьких веревок.
Тут и помог исследователям метод молекулярной селекции. Из библиотеки РНК удалось выделить не-сколько молекул, которые очень успешно связывались с мембранами, а при достаточно высокой концентрации - даже разрывали их! Эти РНК обладали необычными свойствами. Они как бы помогали друг другу: смесь молекул разных сортов связывалась с мембранами гораздо лучше, чем молекулы одного сорта. Все стало ясным после изучения вторичных структур этих РНК. Оказалось, что в них имеются петли с комплементарными участками. За счет этих участков «мембранные» РНК могут формировать комплексы-сообщества, которые способны образовывать множественные контакты с мембраной и делать то, что одной молекуле РНК не под силу.
Этот селекционный эксперимент подсказал, что у РНК есть дополнительный способ приобретения новых свойств путем образования сложных надмолекулярных комплексов. Этот механизм мог использоваться и для удерживания эволюционирующих систем РНК в виде колоний на поверхностях еще до того, как эти системы обзавелись изолирующей мембраной.
«Мир РНК»: Был, Есть и Будет!
Множество данных свидетельствует о том, что «мир РНК» действительно существовал. Правда, не совсем ясно - где. Некоторые специалисты полагают, что начальные этапы эволюции происходили не на Земле, что на Землю были занесены уже функционально активные системы, которые приспособились к местным условиям. Однако с химическойи биологической точки зрения это не меняет сути дела. В любом случае остается загадкой - в результате каких процессов в окружающей среде рибоциты образовались и за счет каких компонентов существовали. Ведь требуемые для жизни рибоцитов нуклеотиды - сложные молекулы. Трудно представить, что эти вещества могли образовываться в условиях пребиотического синтеза.
Вполне возможно, что древние РНК значительно отличались от современных. К сожалению, следов этих древних РНК экспериментально обнаружить нельзя, речь идет о временах, удаленных от нас на миллиарды лет. Даже скалы тех времен давно «рассыпались в песок». Поэтому речь может идти только об экспериментальном моделировании процессов, которые могли протекать на самых ранних стадиях молекулярной эволюции.
Почему произошел переход от «мира РНК» к современному миру? Белки, располагающие гораздо большим набором химических групп, чем РНК, являются лучшими катализаторами и структурными элементами. По-видимому, некоторые древние РНК стали использовать белковые молекулы в качестве «орудий труда». Такие РНК, способные к тому же синтезировать для своих целей полезные молекулы из окружающей среды, получали преимущества в размножении. Естественным путем отбирались соответствующие аптамеры и рибозимы.
А затем эволюция сделала свое дело: возник аппарат трансляции, и постепенно ответственность за катализ перешла к белкам. Орудия оказались столь удобными, что вытеснили своих «хозяев» из многих сфер деятельности.
Читатель вправе спросить: а зачем вообще нужно исследовать эволюцию РНК, ведь древний «мир РНК» исчез? Неужели только ради «чистого искусства», удовлетворения интересов фанатичных исследователей? Однако, не зная прошлого, нельзя понять настоящее. Изучение эволюции и возможностей РНК может подсказать новые направления поиска процессов, протекающих в современных живых клетках. Например, совсем недавно были обнаружены мощные системы регуляции активности генов с участием двуцепочечных РНК, с помощью которых клетка защищает себя от вирусных инфекций. Эта древняя система клеточной защиты, вероятно, скоро найдет применение в терапии.
Поэтому неудивительно, что в наше время исследования нуклеиновых кислот продолжают оставаться одной из самых «горячих точек» в молекулярной биологии. Благодаря уникальным свойствам РНК находят все более широкое применение в медицине и технике. Возникший в незапамятные времена «мир РНК» будет не только продолжать незримо существовать
в наших клетках, но и возрождаться в виде новых биотехнологий.
Редакция благодарит сотрудников Института химической биологии и фундаментальной медицины
СО РАН к. х. н. В. В. Коваля, к. х. н. С. Д. Мызину и к. х. н. А. А. Бондаря за помощь в подготовке статьи
